


植物与土壤微生物之间是相互联系的,作为生产者的植物为土壤微生物提供了碳源,而土壤微生物作为分解者为植物生长提供养分(如图)。同时,由于微生物在分解有机质的过程中只有当自身的营养需求被满足时才会提供植物体养分,所以植物与土壤微生物之间也存在对养分的竞争,植物与土壤微生物间的这种相互依存梁与养分竞争关系在调控生态过程中起着重要作用。
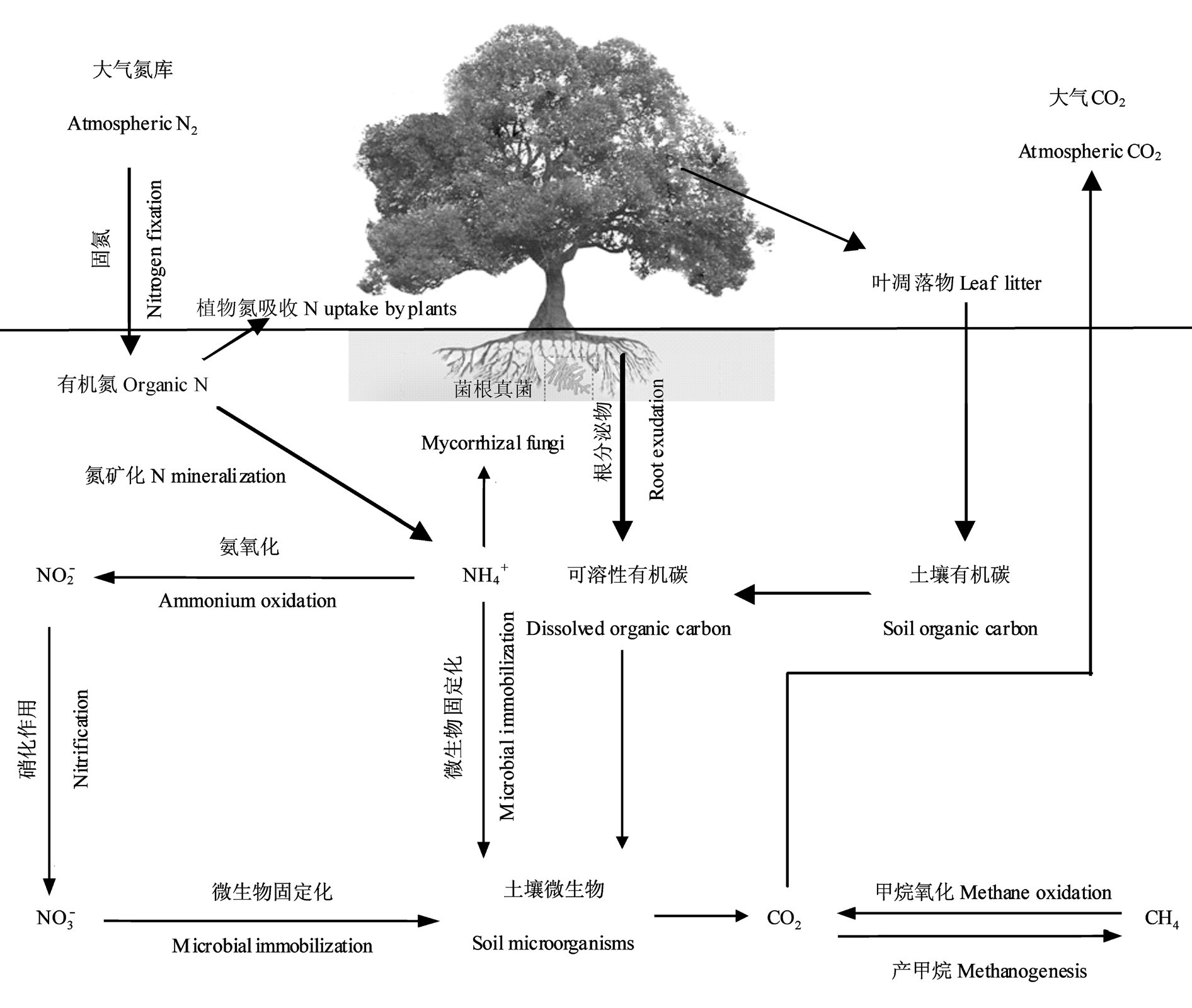
1、植物与土壤微生物之间的相互依存关系
植物通过其凋落物和分泌物为土壤微生物提供营养,导致植物和微生物之间的协同进化,促进土壤微生物的多样性。比如,细菌倾向于利用富含碳水化合物和糖类的凋落物,真菌倾向于利用富含酚类的凋落物;白三叶草根际土壤中的微生物数量和活性与植物根系的长度和密度也高度相关;草地植被生长的根系可增加土壤中原碳的衰变速率。
土壤微生物可以分解可溶性和不溶性有机物,将其转化为植物可以吸收利用的无机形态。微生物固定不可移动氮的季节性格局对植物吸收氮很重要,在氮强烈受限的生态系统,微生物秋季固定的不可移动氮最多,Jaeger等研究了美国科罗拉多州高寒草甸中植物与微生物之间养分的季节性划分,结果表明,优势植物种嵩草(Kobresiamyosuroides)在积雪融化后吸收了大量的氮,而土壤微生物在植物枯黄的秋季固持大量的氮素,并在整个冬季维持这一水平。积雪融化后微生物量氮显著下降,其固持的氮以可溶性氮和有机态氮的形式释放到土壤中,为春季植物的生长提供了养分。与生长季晚期萌发的草本植物功能群相比,生长季早期萌发的植物功能群具有较强的利用早春季节土壤中大量存在的特定形态养分的能力(如在高寒生态系统中生长季早期萌发的植物功能群具有较强的吸收大量存在于土壤中的有机态氮的能力),植物与土壤微生物的生态位分异提高了资源利用的有效性。
此外,植物与土壤微生物共生是自然界中普遍存在的生物学现象。自然群落中90%以上的陆生植物能与泡囊-丛枝菌根真菌(vesicular-arbuscularmycorrhizalfungi,VAMF)共生形成菌根。研究表明,菌根菌在自然界养分循环中的作用,除了能通过根外菌丝将土壤中的矿质元素、水分等输送给植物吸收利用,提高植物成活率,促进植物生长,还能提高植物的抗逆性和抗病性。林鹤鸣等研究表明,在土壤贫瘠的山地条件下,接种外生菌根真菌,可以改善土壤中微生物的种群结构,提高土壤中细菌、真菌、放线菌的数量,其中真菌增加7.3倍,林木的菌根侵染率由20%提高到75%,进而促进油松(Pinustabulaeformis)人工林的生长。当前,国际上有关菌根方面的研究逐渐升温,亦有数篇文章在《Science》、《Nature》等刊物上发表。植物与真菌之间的互利共生关系能提高植物的耐热性。鉴于美国东北部森林生态系统Ca严重流失的情况,Blum等(2002)研究发现外生菌根的树种更能利用有磷灰石风化的Ca,表明菌根可能直接风化磷灰石和吸收释放出的Ca2+,为植物提供钙源。Hodge(2003)发现,在植物种间存在竞争时,接种丛枝菌根可促进植物对氮元素的吸收。Wolfe等(2005)的试验同样表明,在柳兰(Chamerionangustifolium)根部植入丛枝菌根真菌,将大大提高植物被授粉的几率。在养分缺乏的生态系统中,植物共生体吸收限制性养分来调节植物生产力。植物共生体菌根真菌,能增加限制性氮、磷的利用率,对植物生产力有正反馈作用。因此,菌根菌的多样性与丰度对维持植物的多样性及生态系统的稳定性和生产力具有重要意义。
2、植物与土壤微生物对养分的竞争
土壤微生物与植物间的负反馈作用是影响养分循环和群落结构的另一重要因素。植物氮吸收与微生物氮固定向来是研究的热点。在养分强烈受限的生态系统(如北极和高山苔原),微生物与植物在土壤中争夺养分,可能会给植物吸收养分和生长带来负面影响。Xu等观测了青藏高原高寒草甸生态系统植物与土壤微生物对外加氮源的利用情况,结果表明,由于土壤中有效氮匮乏,植物与土壤微生物间存在着对氮素利用的竞争。Song等进一步揭示了植物种间关系调节着植物与土壤微生物间氮素利用的竞争强度。许多研究认为土壤微生物在氮素竞争上具有优势,因为微生物在矿化过程中起主要作用,而且与植物相比,土壤微生物具备特有的属性,如表面积、体积比率大,繁殖速率快等特点,但是也有研究表明,生长在养分受限的区域中的高寒植物具有较强的利用有机氮的能力。
此外,土壤微生物与植物根系的养分竞争以及微生物对养分流失的加速会导致植物生产力的降低。植物与微生物对无机氮(NH4+)的竞争利用可能导致氮硝化基质数量的降低,进而减少了因反硝化和淋溶作用而流失的NO3–的量。病原微生物对植物生产力也有影响,特别是对物种丰富的植物群落。在根围集聚的寄生虫、病原体和根系植食者直接取食植物组织中的碳等营养成分,削弱根部吸收养分的能力,对植物生长产生负反馈作用(Beveretal.,1997)。研究发现,土壤病原菌,如疫霉属(Phytophthora)、镰刀菌属(Fusarium)、腐霉(Pythiumspp.)等对一系列优势树种如橡树(Quercuspalustris)、阿拉伯胶树(Acaciasenegal)、桉树(Eucalyptusspp.)的生长具有抑制作用。
3、植物物种多样性与土壤微生物多样性之间的关系
定居在土壤中的异养微生物群落调节着一些关键的生态过程,这些过程控制着生态系统的C、N循环,它们潜在地体现了植物多样性与生态系统功能之间的一种机理性的联系。很少有研究揭示植物多样性与土壤微生物多样性之间的影响机制。但是,一直以来仍然有很多学者致力于这方面的探索,并有了一定的研究积累。首先,限制植物生长的资源的有效性塑造着生物群落的组成;其次,土壤微生物群落资源的可获取性受到枯死叶和根(凋落物)的化学组分限制,因为凋落物能够被用于产生能量;最后,植物在化学组分上存在差异,植物多样性的改变能够引起植物产物的改变和凋落物有机组分的变化,因此会影响异养微生物群落的组成和功能。Spehn等(2000)的研究表明,土壤微生物数量与植物功能群数量呈线性相关。当功能群中豆科植物缺失时,土壤微生物数量显著降低15%。土壤微生物C与土壤有机C的比值亦随着植物物种的丢失和植物功能群数目的降低而降低。Porazinska等(2003)通过对美国堪萨斯州Konza草原的研究发现,不同组合的C3和C4植物,根系土壤中一些细菌和线虫类对特有植物种反应强烈。有研究表明,植物多样性影响微生物数量主要是通过植物生产力的提高;而另一项研究表明,植物多样性没有影响土壤微生物群落和凋落物的分解。
植物凋落物作为联系地上和地下的桥梁,是研究植物多样性与土壤微生物多样性之间关系的良好媒介。植物凋落物的化学特性较地上植物群落组成、物种丰富度、物种均匀度能更好地解释土壤内部过程。凋落物的质量影响着凋落物中养分和土壤有机质的周转速率,还会对土壤的生物学特征产生强烈影响。有些植物凋落物中含有抑制细菌活动的酚、醛等成分,可间接地影响凋落物的分解率。富含低分子酚类化合物的凋落物会增加所有微生物的生物量,尤其是真菌;而富含碳水化合物和糖类的凋落物会促进细菌的生长。例如,一些生长缓慢的植物(如高寒草本植物Acomastylisrossi)产生的大量的富含酚类的凋落物,进入土壤后控制着真菌占优势的微生物对氮的固持,加剧了低养分的状况;而其他快速生长的植物(如草本植物Deschampsiacaespitosa)表现出较高的细根周转率,生产大量高质量(富含N)的凋落物,促进了细菌占优势的食物网,提高了生境的养分状况。同样,来自适应低养分生境的功能群产生的凋落物分解缓慢,因为低浓度的N、P与高浓度的木质素、丹宁酸、蜡质,以及其他难降解和有毒的化合物对土壤微生物的活性产生了抑制作用,这种负反馈恶化了贫瘠生境中养分的可利用性,反过来会降低植物功能群的生产力。相反,高养分生境中的植物功能群产生易分解的凋落物,从而增强了高养分生境中养分的周转率,提高了植物群落的生产力。总之,植物驱动的基质C、N含量的变化影响着土壤微生物多样性,土壤微生物的活性和生物量以及对氮素的固持,反过来影响着植物多样性。最近《美国国家科学院院刊》上的一项研究结果表明,土壤微生物多样性与作为凋落物的植物化学组分的多样性存在正相关关系,而土壤微生物多样性与植物物种多样性之间不存在直接的相关性。因此,迫切需要通过植物凋落物来建立地上、地下生态过程的联系,从机理上认识植物与微生物之间的作用与反馈。